



Sementara periode dari awal 1900-an Perang Dunia II telah dianggap sebagai "masa keemasan" genetika, para ilmuwan masih belum menetapkan bahwa DNA, dan bukan protein, adalah materi herediter. Namun, selama ini sebuah penemuan besar genetik banyak dibuat dan hubungan antara genetika dan evolusi dibuat.
Friedrich Meischer pada tahun 1869 mengisolasi DNA dari sperma ikan dan nanah dari luka terbuka. Karena berasal dari inti, Meischer menamakan kimia baru, nuklein. Selanjutnya nama ini diubah menjadi asam nukleat dan terakhir menjadi asam deoksiribonukleat (DNA). Robert Feulgen, pada tahun 1914, menemukan bahwa DNA fuchsin bernoda pewarna. DNA kemudian ditemukan dalam inti semua sel eukariotik.
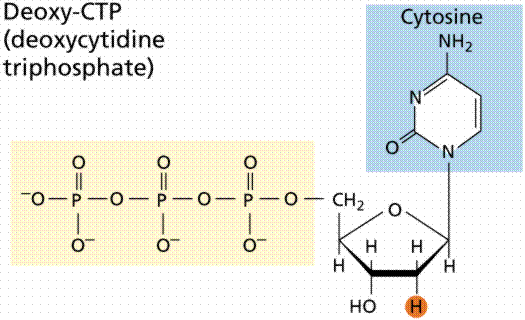
Selama P.A., ahli biokimia 1920 Levene menganalisis komponen dari molekul DNA. Dia menemukan itu berisi empat basa nitrogen: sitosin, timin, adenin, dan guanin, gula deoksiribosa, dan gugus fosfat. Dia menyimpulkan bahwa unit dasar (nukleotida) terdiri dari basis melekat gula dan bahwa fosfat juga melekat pada gula. Dia (sayangnya) juga keliru menyimpulkan bahwa proporsi pangkalan itu adalah sama dan bahwa ada tetranucleotide yang struktur mengulangi molekul. Para nukleotida, bagaimanapun, tetap sebagai unit fundemantal (monomer) dari polimer asam nukleat. Ada empat nukleotida: mereka dengan sitosin (C), mereka dengan guanin (G), mereka dengan adenin (A), dan mereka dengan timin (T).


Molekul struktur tiga basis nirogenous. Dalam diagram ini ada tiga fosfat bukan fosfat tunggal ditemukan di nukleotida normal.
Selama awal 1900-an, studi genetika dimulai dengan sungguh-sungguh: link antara kerja Mendel dan bahwa ahli biologi sel menghasilkan teori warisan kromosom ; Garrod mengusulkan hubungan antara gen dan "kesalahan metabolisme bawaan", dan pertanyaan itu terbentuk : apa itu gen? Jawabannya datang dari studi tentang penyakit menular mematikan: pneumonia. Selama tahun 1920 Frederick Griffith mempelajari perbedaan antara strain penyebab penyakit pneumonia bakteri yang menyebabkan (Streptococcus peumoniae) dan strain yang tidak menyebabkan pneumonia. Strain penyebab pneumonia (strain S) dikelilingi oleh kapsul. Strain lain (galur R) tidak memiliki kapsul dan juga tidak menyebabkan pneumonia. Frederick Griffith (1928) mampu menginduksi strain patogenik dari bakteri Streptococcus pneumoniae menjadi patogen. Griffith disebut faktor transformasi yang menyebabkan bakteri non-patogen menjadi patogen. Griffith menyuntik strain berbeda dari bakteri ke tikus. Strain S membunuh tikus, sedangkan galur R tidak. Dia lebih jauh mencatat bahwa jika panas menewaskan S ketegangan disuntikkan ke tikus, tidak menyebabkan pneumonia. Ketika dikombinasikan panas S yang dibunuh dengan Live R dan disuntikkan campuran ke mouse (ingat tidak saja akan membunuh mouse) bahwa mouse mengembangkan pneumonia dan meninggal. Bakteri pulih dari mouse memiliki kapsul dan membunuh tikus lain saat disuntikkan ke mereka!
Hipotesis:
1. Strain S mati telah reanimated / dibangkitkan.
2. R Hidup telah berubah menjadi S Live beberapa "faktor transformasi".
Percobaan lanjutan dipimpin Griffith untuk menyimpulkan nomor yang 2 itu benar.
Pada tahun 1944, Oswald Avery, Colin MacLeod, dan McCarty percobaan Maclyn revisited Griffith dan menyimpulkan faktor transformasi adalah DNA. Bukti mereka adalah kuat tetapi tidak benar-benar meyakinkan. Favorit kemudian-saat ini untuk materi herediter adalah protein; DNA tidak dianggap oleh banyak ilmuwan menjadi calon kuat.
Terobosan dalam upaya untuk menentukan materi herediter berasal dari karya Max Delbrück dan Salvador Luria pada 1940-an. Bakteriofag adalah sejenis virus yang menyerang bakteri, virus yang Delbrück dan Luria bekerja dengan orang-orang yang menyerang Escherichia coli, bakteri yang ditemukan dalam usus manusia. Bakteriofag terdiri dari mantel protein yang meliputi DNA. Bakteriofag menginfeksi sel dengan menyuntikkan DNA ke dalam sel inang. Ini DNA virus kemudian "menghilang" sementara mengambil alih mesin bakteri dan mulai membuat virus baru, bukan bakteri baru. Setelah 25 menit semburan sel inang, melepaskan ratusan bakteriofag baru. Fag memiliki DNA dan protein, membuat mereka ideal untuk mengatasi sifat materi herediter.

Struktur virus bakteriofag.
Pada tahun 1952, Alfred D. Hershey dan Martha Chase melakukan serangkaian percobaan untuk menentukan apakah protein atau DNA adalah materi herediter. Dengan pelabelan DNA dan protein dengan berbeda (dan saling eksklusif) radioisotop, mereka akan dapat menentukan kimia (DNA atau protein) yang masuk ke bakteri. Bahan tersebut harus menjadi bahan herediter (agen Griffith mengubah). Karena DNA berisi fosfor (P) tapi tidak ada Sulfur (S), mereka ditandai dengan DNA radioaktif fosfor-32. Sebaliknya, protein tidak memiliki P tetapi memiliki S, sehingga dapat ditandai dengan radioaktif Sulfur-35. Hershey dan Chase menemukan bahwa S radioaktif tetap berada di luar sel sedangkan P radioaktif ditemukan di dalam sel, menunjukkan bahwa DNA adalah pembawa fisik keturunan.




Diagram ilustrasi percobaan Hershey dan Chase yang mendukung DNA sebagai materi herediter sementara itu juga menunjukkan protein TIDAK materi herediter.
Struktur DNA
Erwin Chargaff menganalisis basa nitrogen dalam berbagai bentuk kehidupan, menyimpulkan bahwa jumlah purin tidak selalu sama dengan jumlah pirimidin (seperti yang diusulkan oleh Levene). DNA telah terbukti sebagai materi genetik oleh Hershey-Chase percobaan, tetapi bagaimana DNA menjabat sebagai gen itu belum bisa dipastikan. DNA harus membawa informasi dari sel induk untuk sel anak. Ini harus berisi informasi untuk mereplikasi dirinya sendiri. Pasti kimiawi stabil, relatif tidak berubah. Namun, harus mampu perubahan mutasi. Tanpa mutasi tidak akan ada proses evolusi.
Banyak ilmuwan tertarik untuk menguraikan struktur DNA, di antara mereka adalah Francis Crick, James Watson, Rosalind Franklin, dan Maurice Wilkens. Watson dan Crick mengumpulkan semua data yang tersedia dalam upaya untuk mengembangkan model struktur DNA. Franklin mengambil photomicrographs difraksi sinar-X ekstrak DNA kristal, kunci teka-teki. Data yang dikenal pada saat itu adalah bahwa DNA merupakan molekul panjang, protein yang spiral melingkar (sebagaimana ditentukan oleh karya Linus Pauling), basis data Chargaff, dan data difraksi sinar-x dari Franklin dan Wilkens.

DNA adalah double heliks , dengan basis ke pusat (seperti anak tangga pada tangga) dan gula-fosfat unit sepanjang sisi helix (seperti sisi tangga pair). Untaian saling melengkapi (dideduksi oleh Watson dan Crick dari data Chargaff, A berpasangan dengan T dan C dengan G pasangan, pasangan yang diselenggarakan bersama oleh ikatan hidrogen). Perhatikan bahwa purin ganda bercincin selalu terikat pada cincin pirimidin tunggal. Purin adalah Adenin (A) dan Guanin (G). Kami mengalami Adenosin trifosfat (ATP) sebelumnya, meskipun dalam hal ini gula adalah ribosa, sedangkan pada DNA adalah deoksiribosa. Pirimidin adalah Sitosin (C) dan Timin (T). Dasar saling melengkapi, dengan A pada satu sisi dari molekul Anda hanya mendapatkan T di sisi lain, sama dengan G dan C. Jika kita mengetahui urutan dasar satu untai kita tahu komplemennya.


Model pita DNA
Replikasi DNA
DNA terbukti sebagai materi herediter dan Watson et al. telah diuraikan strukturnya. Apa yang tersisa adalah untuk menentukan bagaimana DNA disalin informasi dan bagaimana yang dalam phenotype. Matius Meselson dan Franklin W. Stahl dirancang percobaan untuk menentukan metode replikasi DNA. Tiga model replikasi dianggap mungkin.
1. Replikasi konservatif entah bagaimana akan menghasilkan untai DNA yang sama baru sekali selama replikasi.

2. Replikasi semikonservatif akan menghasilkan dua molekul DNA, yang masing-masing terdiri dari satu-setengah dari DNA induk bersama dengan untai komplementer sama baru sekali . Dengan kata lain DNA baru akan terdiri dari satu untai tua baru dan satu DNA. Alur yang ada akan berfungsi sebagai template untuk untai komplementer baru.

3. Replikasi dispersif terlibat pemecahan dari untaian induk selama replikasi, dan entah bagaimana, sebuah molekul yang reassembly adalah campuran fragmen lama dan baru pada setiap untai DNA.

Percobaan Meselson-Stahl terlibat pertumbuhan bakteri E. coli pada medium pertumbuhan yang mengandung nitrogen berat (Nitrogen-15 yang bertentangan dengan isotop berat lebih umum, tetapi lebih ringan molekul, Nitrogen-14). Generasi pertama dari bakteri ditumbuhkan pada media mana satu-satunya sumber N adalah Nitrogen-15. Bakteri kemudian dipindahkan ke media dengan cahaya (Nitrogen-14) menengah. Watson dan Crick telah meramalkan bahwa replikasi DNA adalah semi-konservatif. Jika itu, maka DNA yang dihasilkan oleh bakteri ditanam pada medium cahaya akan menjadi penengah antara berat dan ringan. Itu.
Replikasi DNA melibatkan banyak blok bangunan besar , enzim dan banyak ATP energi (ingat bahwa setelah fase S dari siklus sel sel memiliki fase G untuk menumbuhkan energi untuk pembelahan sel). Hanya terjadi dalam sel sekali per (sel) generasi, replikasi DNA pada manusia terjadi pada tingkat 50 nukleotida per detik, 500/second di prokariota. Nukleotida harus dirakit dan tersedia dalam inti, bersama dengan energi untuk membuat ikatan antara nukleotida. DNA polimerase unzip helix dengan memecah H-ikatan antara basa. Setelah polimerase telah membuka molekul, sebuah daerah yang dikenal sebagai bentuk replikasi gelembung (selalu dimulai pada satu set tertentu dari nukleotida, asal replikasi). Nukleotida baru ditempatkan di garpu dan link ke orangtua nukleotida yang sesuai sudah ada (A dengan T, C dengan G). Prokariota membuka gelembung replikasi tunggal, sedangkan eukariota memiliki gelembung ganda. Seluruh panjang dari molekul DNA direplikasi sebagai bertemu gelembung .


Peran polimerase DNA dalam replikasi DNA.
Karena untai DNA adalah hasil antiparalel, dan replikasi di thje 5 'ke 3' arah pada SETIAP untai , satu untai akan membentuk salinan terus menerus, sementara yang lain akan membentuk serangkaian fragmen Okazaki pendek.



semoga bermanfaat ^ ^







0 komentar:
Posting Komentar